|
SIZE INHERITANCE IN THE CARDIGAN WELSH CORGI One
of the most striking features
of the domestic dog is its enormous variety of size and shape, not found
in any other species, either wild or domestic one. The cardigan corgi is
an example of a short-legged, long bodied (chondrodysplastic)
breed. Research suggest that a single mutation appeared
at very early stage in domestic dogs development, and it is commonly
found in small breeds. Parker
et al. (2009), using multibreed association analysis in domestic dogs,
demonstrate that expression of a recently acquired retrogene encoding
fibroblast growth factor 4 (fgf4) is strongly associated with
chondrodysplasia, a short-legged phenotype that defines at least 19 dog
breeds, including dachshund, corgi and basset hound. Size,
together with many other features, that contribute to dog appearance, is
a polygenic, quantitative trait . This means that it is measureable (e.g.
size, or height at withers in cms, body weight in kgs), and variable
within the breed. These
traits depend mostly on additive influence of many (the exact number is
not known) different genes, and are
also influenced by environment. In case of size, the final result
depends both on individual genetic makeup and proper rearing.
Recent,
more advanced studies, employing sophisticated methods of molecular
genetics, identified 6 loci
of quantitative genes (QTLs) that influence size and shape in dogs (Chase
et al.2002). Mutations within
IGF-1 (insulin-like growth factor 1) were found
of crucial importance during
puppy growth; they also influence metabolism in adult dogs
(Sutter et al. 2007) . It was also shown
that small and miniature breeds are usually homozygous in three
IGF-1 haplotypes.
Specimens
of any breed vary in size, and this variation is usually included in
their respective breed standards. In case of cardigan welsh corgi the
standard reads: “ideal
height at withers 30 cms”. However, in any group of cardis there will
be probably no two dogs of exactly same size.
Where does this variation come from (apart from different rearing
conditions)? To begin, assume that all cardigans have same genes,
encoding the actual size, and their accumulated action enable any given
specimen to grow to 30 cms. The only difference is in two pairs of
additive alleles, A1A2 and B1B2. Assume also that A1 and B1 add 1 cm to
the actual height, whereas A2 and B2 do not increase the size.
When two specimen of identical heterozygous
genotypes are mated
together, there will be 16
possible combinations of gametes (see the table) : TABLE 1:
In resulting progeny there may be:
1/16 puppies of 34 cms height
- 6.25% The
majority of puppies will be these of medium height (same as their
parents); however, there will be also several specimens distinctly
smaller or
bigger than parents. It
is important to realize that the above example is most probably
simplified. Variation in size results not only from additive action of
genes. In some cases there might be also two other types of co-operation
within or between alleles
involved. Assume that in the given pair of alleles
- A1A2 - A1 is
completely dominant to A2. In this case a dog of A1A1 genotype does not
differ in size to a dog of A1A2 (in our example this will give at least
32 cms, the rest depending on locus B). Yet the first dog (A1A1) would
pass the “big size” gene to all its puppies, whereas the other dog (A1A2) – only to half of them. Additionally,
genes of one allele pair may be epistatic to genes of the
other allele. An epistatic gene (or its pair)
blocks other gene; in our example it may happen that A2A2 pair
block completely the action of B1 gene… Then a dog of A2A2B1B1
genotype is only 30 cms
tall, and not 32 cms. As far as size is concerned, it is possible that
the presence of certain mutated IGF
1 allele, which results in low production of insulin,
leads itself to small size, even though the combination of other
genes could bring bigger size. Epistatic co-operation between different
genes is not hereditary: A2A2B1B1 dog will pass B1 and A2 alleles to its
puppies; if they get A1
allele from the second parent, epistatic effect will not appear.
To
make the matter even more complicated, quantitative traits are – as
written earlier - influenced
by environment. This influence (optimal
or less favorable conditions during growth period) may result in
increase or decrease of the actual height, let say by
+0.3 to -0.3 cm. Puppies, which are genetically 34 cms, will show
slightly different size – from 33.7 up to 34.3 cms, depending on
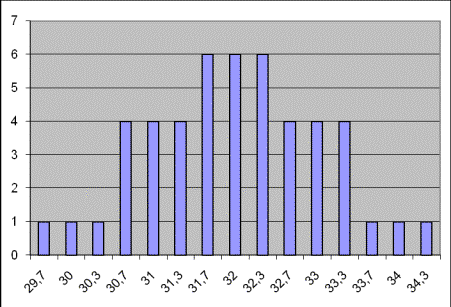
whether they have been reared in excellent, decent, or poor conditions. Graph
1 shows size variation in any given group of cardigan welsh corgis: Graph
1
As
it is seen on the graph, size variation varies from 29.7 to 34.3 cms,
and the majority of dogs remains within medium size range. As the
number of genes, contributing to size, is unknown, it is worth
remembering that the number of possible genotypes may be enormous. In
case of only one pair of alleles there are 3 possible genotypes, in case
of two pairs – 9 different genotypes, in case of n pairs
- 3n different
genotypes. Just as few as 6 pairs of alleles result
in 36 = 729 different genotypes possible. Considering
additional co-operation of genes, as described earlier, and
environmental influences, it does not come as a big surprise that the
number of possible phenotypes is almost endless, and they might be very
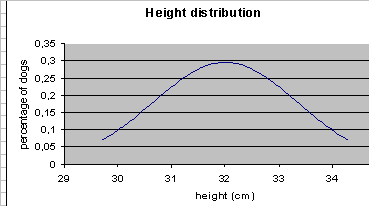
close to each other. They
cannot be shown on the graph like Graph 1,
for they form continuum, bordered by two values – the highest
and the lowest one, as shown on Graph 2. GRAPH 2.
Typically,
variability of any quantitative trait can be graphically presented in
the form of bell- shaped curve (Gauss curve) with its peak at mean value.
This type of distribution is known as “normal”. It can be seen that
the majority of dogs, i.e. 1/3 of the total population, are these of
medium size – 32 cms. Size of appr. 68% of all dogs remains within standard
deviation (SD) range from the mean value (in our case – 32 cms), and
98% - within double SD range. This is
only an example – the actual dispersion, or variability in the
population can be bigger or smaller, resulting in more flat or more
steep curve. In breeding farm animals selection may be aimed at increasing size and weight (e. g. poultry, swine).
In pure bred dogs, with just a few exceptions,
such type of selection does not take place, at least consciously,
and probably no one has ever selected cardigans for their size only. Nevertheless, they definitely
have become bigger, and
one may ask the reason. There
may be at least three reasons that lead to increase in cardigan size: 1.
Improved environmental conditions, mainly balanced feeding, that enables
a dog to fully develop its
genetic potentials. 2.
Unconscious selection. Very few breed standards call for eliminating from
breeding any specimen above or under specified height. In Shetland
Sheepdogs breeders avoid using dogs that are obviously oversized, as
height of more than +2.5 cms above the standard
is considered a serious fault (at least in GB and Europe).
No cardigan breeder will probably avoid breeding oversized
animals; moreover, in practice such specimens are favored in
conformation rings as they look showy. Contrary, a small specimen,
especially a male, will be probably faulted (or at least ignored by
judges) and thus not used, or seldom used in breeding. In our simplified
example (Table 1) an A2A2B2B2
male will be the one which is not used, whereas A1A1B1B1 will be
probably used frequently.
By culling smallest specimens the mean height of dogs in
subsequent generation will be slightly bigger than that of parental
generation. In our example, if
all dogs which are smaller than 30 cms are not used in breeding,
the mean height of parental generation will increase to 32.3 cms. Such
“unconscious” selection has low intensity (especially considering
that size itself is seldom the main factor in selection) , yet it
accumulates in subsequent generations and is growing slowly, but
steadily, and the mean height of the population gradually
increases. (N.B:
The actual increase depends also on
heritability (h2) of
the trait. This is one of the most important parameters used
in scientific breeding,
and indicates what part of the total variability in phenotypes is
inherited. h2
of any given trait can vary
from 0 to 1, or 100% –
the higher it is, the bigger is genetic
compound of the trait). 3.
It is commonly known that small dogs produce smaller litters than dogs of
large breeds (Willis, 1989). Also in one breed, e.g. Yorkshire Terrier,
small bitches deliver small litters. Thus small cardigan bitches may
produce smaller litters and introduce
fewer puppies to the
next generation than bigger
bitches. In the next generation there will be more specimens of bigger
size and carrying such genotype. (revised by Katarzyna
Fiszdon, PhD, Dept. Of Animal Breeding, Warsaw University of Life
Sciences) Literature: Parker
H.G. et al. An Expressed fgf4 Retrogene Is Associated with
Breed-Defining Chondrodysplasia in Domestic Dogs, Science: Vol. 325
(2009), pp. 995 – 998
|
|||||||||||||||||||||||||||